BULLETIN DE L'INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE.
SCIENCES DE LA TERRE, 66-SUPP.:95-102, 1996
BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETTENSCHAPPEN
AARDWETENSCHAPPEN, 66-SUPP.: 95-102, 1996
_____________________________________________________________
with contributions by Jackie A. Burnett, Christopher J. Collom, Walter K. Christensen, Annie V. Dhondt, Silvia Gardin, Kai-Uwe Gräfe, Jana Ion, Erle G. Kauffman, William James Kennedy, Ludmila F. Kopaevich, Gregorio López, Tatsuro Matsumoto, Rory Mortimore, Isabella Premoli-Silva, Francis Robaszynski, Jozef Salaj, Herbert Summesberger, Seichii Toshimitsu, Karl-Armin Tröger, Michael Wagreich, Christopher J. Wood, Elena A. Yazykova.
The recognition of the Coniacian-Santonian boundary is easy with good correlation of macro- and microfossil evidence. The Santonian Working Group (SWG) recommends the lowest occurrence of Cladoceramus undulatoplicatus (Roemer) as the marker for the Coniacian-Santonian boundary. As yet, the SWG cannot make a formal proposal for a Boundary Stratotype Section, because the biostratigraphy must be better known and integrate first. Three candidates for Boundary Stratotype Section, Olazagutia Quarry (Navarra, Spain), Seaford Head (Sussex, England) and Ten Mile Creek (Dallas, Texas, USA), were selected for further decision. To achieve an useful subdivision of the Santonian into sub-stages a better understanding of taxa ranges and correlation through different biogeographic realms is needed. Formal proposals for subdivision would be premature at present, but a three-fold division is favoured.
Keywords: Santonian, Upper Cretaceous, stratotypes, chronostratigraphy, biostratigraphy, ammonites, belemnites, crinoids, foraminifera, inoceramids, nannofossils.
The Santonian Stage was proposed by Coquand (1857), presumably named after the town of Saintes in south-west France. One of the localities which Coquand mentioned was Javrezac, a village on the north-west side of Cognac. The boundary there was drawn on a hardground between glauconitic, nodular limestone, with many Exogyra of the Coniacian below, and soft micaceous chalk of the Santonian above.
At the First Symposium on Cretaceous Stage Boundaries (Birkelund et al. 1984) the consensus was that the first appearance of Texanites (Texanites) and of Cladoceramus undulatoplicatus (Roemer) are the two best boundary criteria. Texanites (Texanites) has been used over a wide area, although in classic regions of north-west Europe this subgenus is far too rare to be a practical marker (Hancock, 1991). Cladoceramus undulatoplicatus is widespread, and owing to its characteristic form and sculture is easy to determine. The appearance of Platyceramus siccensis (Pervinquière) is well known in N. Africa associated with Texanites, and it could be another possible marker for the base of the Santonian.
In the Santonian Working Group report to the IInd Symposium on Cretaceous Stage Boundaries other macro- and microfossils were mentioned as possible indices for the Coniacian-Santonian Boundary.
Thus, the lowest occurrence of the ammonite Placenticeras polyopsis (Dujardin) is relatively close to lower Texanites (Texanites). Similarly, the entry of the Sphenoceramus pachti-cardissoides group is a possible marker for this boundary, associated with Texanites and Cladoceramus undulatoplicatus. Further discussion on macrofossil criteria were reported by W.J. Kennedy (1995).
Published calcareous nannofossil events do not precisely define the Coniacian-Santonian boundary, which Sissingh (1977) placed in CC14 (above the first occurrence (F.O.) of Micula staurophora (Gardet) Stradner); e.g., lower occurrence of Reinhardtites anthophorus (Deflandre) Perch-Nielsen is unreliable (Wagreich, 1992), and recent studies (e.g., at Ten Mile Creek; Burnett, written comm.) show, however, that this boundary actually lies above the F.O. of Reinhardtites anthophorus (i.e. in CC15 of Sissingh, 1977). Nannofossil event which approximate the boundary are (from stratigraphically old to young): F.O. of Lithrastrinus grillii Stradner, last occurrence (L.O.) of Quadrum gartneri Prins & Perch-Nielsen, L.O. of Flabellites oblongus (Bukry) Crux (all below the boundary) and the L.O. of Lithrastrinus septenarius Forchheimer above the boundary. All these events fall within CC15. Other nannofossil events, such as the L.O. of Eprolithus floralis (Stradner) Stover, and F.O. of Micula concava (Stradner) Verbeek occur later or earlier than entries of Texanites and Cladoceramus undulatoplicatus. Nevertheless, Flores et al. (1987) commented that Micula concava and Lithrastrinus grillii are present in greater abundances in the Santonian than in the Coniacian, and this may serve as a general guide, although nannofossil abundance is related to ecological and taphonomical factors which are generally not cosmopolitan in their extent.
First occurrences of the planktonic foraminifera Sigalia deflaensis (Sigal) and Sigalia carpatica Salaj have been used as markers of the latest Coniacian and the Coniacian-Santonian Boundary (Salaj, 1975; Sigal, 1977, respectively). Also, the first occurrence of Dicarinella asymetrica (Sigal), belonging to Dicarinella concavata group, dates early Santonian, since it post-dated lower Texanites and Cladoceramus undulatoplicatus. Successive occurrences of these three taxa are very promising for correlation with macrofaunal ranges, and a definition of the Coniacian-Santonian Boundary.
Boundary criteria proposed at Copenhagen, and other ones commented in the Working Group reports, were discussed at Brussels during the 2nd Symposium in 1995.![]()
Once it had been concluded that the species Texanites texanus Roemer does not occur in the Aquitaine Basin, nor in other European localities, (Birkelund et al., 1984) pointed out the first appearance of the subgenus Texanites (Texanites) could still be a good marker of the Coniacian-Santonian boundary. Nevertheless, Texanites (Texanites) is rare in many areas where other indices occur, only in Zululand (South Africa) and Madagascar is the subgenus common (reported by Kennedy, 1995).
In the Anglo-Paris Basin, Texanitidae are rare, thus in a hard-ground level in the Craie de Villedieu, France, Texanites gallicus Collignon occurs with Placenticeras polyopsis Dujardin and Inoceramus (Cladoceramus) (Jarvis & Gale, 1984). In S. England, Sussex, Texanites has not been found, but Spinaptychus, an aptychus associate with Texanites, is recorded below and close to the Cladoceramus undulatoplicatus event (Bailey et al., 1983, p. 34; see below inoceramid chapter).
Kennedy (1995) cited Germany as a region where there are well documented co-occurrences of Texanites and Cladoceramus undulatoplicatus (Roemer), particularly the species Texanites (Texanites) pseudotexanus de Groussouvre (=Texanites texanus auct.). Seitz (1961) cited in the shaft Ewald Section the coexistence of Texanites texanum (= Texanites (T.) pseudotexanus) and Inoceramus pachti (Arkhanguelsky) (fide Tröger, written comm.; see below inoceramid chapter).
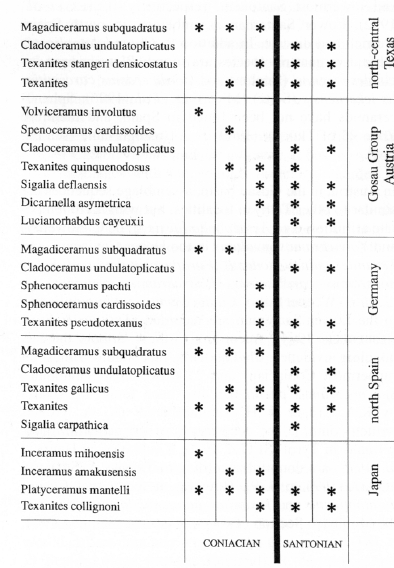
In the Gosau Group, in Austria, (Tröger and Summesberger, 1994), Texanites quinquenodosus (Redtenbacher) co-occurs with Sphenoceramus cardissoides Goldfuss and Parapuzosia daubreei (De Groussouvre). This assemblage is several metres below the first occurrence of C. undulatoplicatus, and above the Coniacian inoceramid Volviceramus involutus (Sowerby). Such a sequence is well correlated with Tröger's (1989) inoceramid biostratigraphy. Wagreich (1992) correlated lower Santonian localities in Austria (Gosau Valley and Bad Ischl-Nussensee) with Texanites quinquenodosus, asociated with the first occurrences of the planktonic foraminifera Sigalia deflaensis and Dicarinella asymetrica. Also, those assemblages were directely correlated with the Cladoceramus undulatoplicatus-bed in the Brandenberg area.
In the Pyrenees Texanites (Texanites) occurs at several levels in the Santonian. In N. Spain (Martínez et al., 1996) Texanites (T.) hispanicus Collignon, T. cf. quinquenodosus, T. gallicus) occurs in several sections. These species have an irregular distribution, but Texanites gallicus and Texanites sp. are sometimes found below C. undulatoplicatus, more than 40m in the Oteo section (López, written comm.). In Tunisia, also a Tethyan area, at Djebl Fguira Salah Texanites (T,) olivetti (=T. quinquenodosus) occurs in the lowermost Santonian levels, with Platyceramus siccensis (Pervinquière) (fide Birkelund et al., 1984; Salaj, 1980, p. 91). In the Corbières, S.E. France, Texanites soutoni (Baily), Texanites quinquenodosus and T. gallicus occur in "Middle" Santonian but not in in the Lower Santonian Nowakites carezi Subzone (Kennedy et al., in press); as in the Craie de Villedieu, T. gallicus co-occurs with P. polyopsis. Although no texanitids have been found above a distinctive Upper Coniacian Paratexanites serratomarginatus Zone, with Protexanites spp., there is otherwise a macrofaunal turnover, whose species belong to the genera Muniericeras, Texasia, Pseudoschloenbachia, Nowakites, etc., which mark the base of the Santonian (Kennedy et al., in press).
Near Austin in Texas the Texanites stangeri densicostatus Zone, as understood by Keith Young, begins some distance (but probably 2 m) below the lowest C. undulatoplicatus (Young, 1963, particularly fig. 4); moreover, its main abundance is near the top of the Texanites stangeri densicostatus Zone. In the Dallas area of north central Texas there is a record of Texanites (Larson et al., 1991, fig. 10.6) in a bed which A. Gale and J. Hancock found to be at least 12m below the lowest C. undulatoplicatus (Hancock, written comm.). This figure agrees with data from N. Spain. Therefore the lowest Texanites cannot be used as an accurate and absolute standard for the base of the Santonian stage.
At Amakusa, in S.W. Japan, the Coniacian-Santonian boundary is marked by the occurrence of Texanites collignoni Klinger and Kennedy (=Texanites olivetti auct.) which is associated with Inoceramus amakusensis Nagao and Matsumoto (Ueda, 1962; Matsumoto, written comm.), whereas in the Haboro area, N.W. Hokkaido, I. amakusensis occurs below T. collignoni (Toshimitsu et al., 1995, fig. 2, p. 24), but just above the Coniacian species Inoceramus mihoensis Matsumoto.
The first appearance of Texanites, particularlyT. (Plesiotexanites) kawasakii (Kawada) and I. amakusensis are the two best criteria for the Coniacian-Santonian boundary in Sakhalin. Both species lie above Peroniceras sp. and I. mihoensis (Yazykova, written comm., Yazykova, 1996).![]()
Three species have been used to define the Coniacian-Santonian boundary: the widespread species Cladoceramus undulatoplicatus, the Northern Temperate Sphenoceramus pachti-cardissoides group and the N. African Platyceramus siccensis.
The last species, possibly related to the Platyceramus cycloides group, was cited in Algeria (Sigal, 1952) and Tunisia associated with Sigalia carpatica Salaj; both occurrences are above the late Coniacian species Sigalia deflaensis (Sigal) (Salaj, 1980). No occurrence is known outside N. Africa.
Cladoceramus undulatoplicatus is a worldwide species, which has been used to define the Coniacian-Santonian boundary. It is asociated with Sphenoceramus in North Temperate areas, and lies above Magadiceramus subquadratus (Schlüter) in the Northern Hemisphere (Dhondt, 1992; López et al., 1992).
In the Western Interior of N. America, Kauffman et al.(1993) reported the co-occurrence of Cladoceramus undulatoplicatus s.s., Sphenoceramus pachti pachti? and Platyceramus cycloides (Wegner) at the base of the Santonian, which are asociated with Cordiceramus cordiformis (Sowerby) in overlying beds. Near Austin, Texas, the lowest Cladoceramus undulatoplicatus occurs at an uncertain distance (but probably about 2 m) above the base of the Zone of Texanites stangeri densicostatus (see above, ammonite chapter). In the Dallas area of north central Texas there are a volvoceramid assemblage and specimens of Texanites overlain by C. undulatoplicatus in a bed which Andy Gale and Jake Hancock found to be at least 12 m above (Hancock, written comm.)
In the North German-Polish Basin it is common to find the succession of uppermost Coniacian Magadiceramus subquadratus (Tröger's Zone 24) and lowermost Santonian Sphenoceramus pachti and Sphenoceramus cardissoides (Tröger's Zone 25). They are followed (Tröger, 1989) by occurrences of Cladoceramus undulatoplicatus. Seitz (1961, Abb. 2, p.19) cited, in the shaft Ewald section, the coexistence of Sphenoceramus pachti and Texanites texanum (Roemer) (=Texanites (T.) pseudotexanus), and Seitz (1965) mentioned occurrences of Cladoceramus undulatoplicatus cf. michaeli (Heinz) with Texanites sp.; he also cited the sequence of Sphenoceramus cardissoides subsp. ind. and S. pachti followed by Texanites pseudotexanus at Recklinghausen (fide Tröger, written comm.).
In England there is a double Cladoceramus event, which can be traced throughout the Anglo-Paris Basin, N.W. Germany (Bailey et al., 1984) and N. Spain. Below these levels, in S. England there are no occurrences of sphenoceramids: the lower occurrence of Sphenoceramus cardissoides was collected above the double Cladoceramus event (Bailey et al., 1983; Wood, written comm.) Recent field work by A. Gale in the Isle of Thanet (S.E. England) has demonstrated that in addition to the two main Cladoceramus events, there are three additional minor Cladoceramus occurrences in the overlying succession. This total known range of Cladoceramus matches occurrences of Spinaptychus. There is no evidence in S. England for the existence of a Sphenoceramus pachti-cardissoides group Zone below the Cladoceramus undulatoplicatus Zone (Wood, written comm.).
In both France and N. Spain, inoceramid assemblages are good markers for the Coniacian-Santonian boundary. Magadiceramus subquadratus Schlüter followed by Cladoceramus undulatoplicatus characterize the uppermost Coniacian and lowermost Santonian, respectively (López et al., 1992). Lower Santonian assemblages, in N. Spain, are rich and diverse. In addition to C. undulatoplicatus, other divergent rib species are found, Platyceramus cycloides wegneri Böhm and Cordiceramus cordiinitialis ickernensis Seitz (Martínez et al., 1996). Sphenoceramids have not been found in Spain, and therefore Zone 25 of Tröger (1989) is not recognisable (see below).
In Austria, in the Gosau basin, assemblage sequences are similar to other Tethyan localities, but with north temperate affinities, e.g., Actaeonella laevis (Sowerby) and Volviceramus involutus (Sowerby) in the Upper Coniacian, and Texanites quinquenodosus, Parapusozia daubreei, Sphenoceramus cardissoides, Platyceramus cycloides cycloides (Wegner) and Cladoceramus undulatoplicatus in the Lower Santonian are recorded (Tröger and Summesberger, 1994). According to these authors there is no clear evidence of Tröger's inoceramid zones 24 (uppermost Coniacian) and 25 (lowermost Santonian). S. pachti have been also found in Austria by K.A. Tröger and H. Summesberger (Tröger, written comm.). Occurrences of the genus Magadiceramus in Tyrol (Seitz, 1970; Kauffman, in Herm et al., 1979) are doubtful after Tröger and Summesberger (1994, p. 179). However, it seems that Cladoceramus undulatoplicatus and Texanites quinquenodosus co-occur in the Mühlbach Section (Tröger and Summesberger, 1994: 184).
In the Caucasus Region and Central Asia (Turkmenia; Moskvin, 1986) the Coniacian-Santonian transition shows an upper Coniacian with Volviceramus involutus overlain by Cladoceramus undulatoplicatus, and sometimes Sphenoceramus cardissoides,
In far-east Asia (Japan, Sakhalin) Inoceramus amakusensis Nagao and Matsumoto is a well known marker for the base of the Santonian, associated with Texanites (T.) collignoni or Texanites (Plesiotexanites) kawasakii (Kawada). The succession of Inoceramus mihoensis overlain by I. amakusensis characterizes the Coniacian-Santonian Boundary in the Haboro area (Hokkaido; Toshimitsu et al., 1995), and Sakhalin (Yazykova, written comm.). Some Coniacian inoceramid species, in Europe, may have higher occurrences in far-east Asia, since Platyceramus mantelli (de Mercey) is known in Upper Coniacian and also "Santonian" together with I. amakusensis (Matsumoto, written comm.). The species Cladoceramus undulatoplicatus is unknown in far-east Russia.
Sphenoceramids are widespread except for rare occurrences in the Tethyan Realm. The species Sphenoceramus pachti and S. cardissoides characterize the lowermost Santonian Tröger's Zone 25, but this zone is only well recognized in the North German-Polish Basin, where both species are associated with Sphenoceramus? bornholmensis Tröger & Christensen, and Platyceramus cycloides ahsenensis Seitz (Tröger, 1989). Nonetheless, either S. pachti and S. cardissoides, mainly the latter, are recorded in many localities from the Northern Temperate Realm (Dhondt, 1992; López et al., 1992), overlying Coniacian volvoceramids or Magadiceramus assemblages, and underlying Cladoceramus undulatoplicatus and Platyceramus cycloides wegneri Böhm assemblages.
Even in the Gosau Basin, a tethyan locality, S. cardissoides is recorded close to Cladoceramus undulatoplicatus (Tröger and Summesberger, 1994), but probably underlying it. Sphenoceramids have also been recorded in the Charente, S.W. France (Dhondt, 1992), on a northern border of the Tethyan Realm. In contrast, in other Tethyan localities, e.g., Crimea and Caucasus (Dobrov and Pavlova, 1959), S. cardissoides is associated with C. undulatoplicatus.
Up till now, no Sphenoceramids have been found in N. Spain, but the especies Platyceramus cycloides ahsenensis is common and is associated with C. undulatoplicatus, as in the N. German-Polish Basin. In consequence, it is possible to make a good correlation between Lower Santonian rocks belonging to the Northern Temperate and Tethyan Realms (see López et al., 1992, for additional common species in N. Germany and N. Spain) (Figure 1).![]()
This group shows different assemblages controlled by biogeography. In general, low and middle latitudes have good occurrences of planktonic foraminifera, whereas in the Temperate Realm benthic foraminiferal assemblages characterize the Coniacian-Santonian boundary.
In the Mediterranean area of the Tethys, the succession of species Sigalia deflaensis (Sigal), Sigalia carpatica Salaj and Dicarinella asymetrica (Sigal) characterize the Coniacian-Santonian transition. Sigalia deflaensis has a widespread occurrence in the Tethyan Realm (see Master, 1977), but S. carpatica is only known from the Mediterranean Region and, like D. asymetrica, was also rare in shallow water facies. Nevertheless, as the lower occurrence of S. carpatica is associated with C. undulatoplicatus and Texanites (Texanites) spp. in N. Spain (Martinez et al., 1996), or with Platyceramus siccensis in N. Africa (Salaj, 1980), it is a good regional index (Sigal, 1977) which should be checked throughout the Tethyan Realm, where both Sigalia deflaensis and D. asymetrica associated with inoceramids and texanitids occur. Both species S. deflaensis(=Sigalia carpatica; Wagreich, written comm.) and D. asymetrica, with D. concavata (Brotzen), were cited by Wagreich (1992) in Austria, as markers of the Coniacian-Santonian Boundary. Especially, in the Bad Ischl-Nussensee section successive occurrences of S. deflaensis, D. asymetrica and Sigalia decoratissima (de Klasz) are similar to Tunisian localities.
Nannofossil occurrences in these levels are similar to other boundary sections, e.g., rare upper Eprolithus floralis (Stradner) Stover, Lithrastrinus septenarius Forchheimer and Quadrum gartneri Prins & Perch-Nielsen, associated with lower occurrences of Lithrastrinus grillii Stradner and Micula decussata Vekshina (=Micula staurophora (Gardet) Stradner), in upper Coniacian beds. All these entries lie below Lucianorhabdus cayeuxii Deflandre (Wagreich, 1992), an early Santonian species.
The species Stensioeina polonica Witwicka used to be a good marker of the Coniacian-Santonian boundary in the North Temperate Region, from England to East Europe. In S.E. England Stensioeina polonica occurs with lower C, undulatoplicatus, which marks a significant macrofaunal and microfaunal turnover (Wood, written comm.). In Eastern Boreal Europe the F.O. of Stensioeina exsculpta exsculpta (Reuss) is a very convenient marker for the Coniacian-Santonian boundary ((Kopaevich, written comm.). Other benthic species, such as Neoflabellina gibbera (Wedekind), occur in N. Africa and Western Carpathia, and its stratigraphical range is similar to Sigalia carpatica (Salaj, 1980). Furthermore, this author correlated the Sigalia carpatica Zone from W. Carpathia with the lower part of the Anomalina (Anomalina) infrasantonica Zone (Vasilenko, 1961), allowing a correlation of Western Tethys with its Eastern European areas.
1) F.O. of Texanites (Texanites)
2) F.O. of Sigalia carpatica
3) F.O. of Dicarinella asymetrica
4) F.O. of Platyceramus siccensis
5) F.O. of Cladoceramus undulatoplicatus
6) F.O. of Sphenoceramus pachti
No nannofossil event is suitable for the Coniacian-Santonian boundary
Selected marker for the Coniacian-Santonian boundary
Primary marker: The lowest occurrence of Cladoceramus undulatoplicatus (Roemer). It is a taxon easely recognisable and widespread. It is known from N. America, Europe, Africa, Madagascar, and Central Asia.
This proposal was supported by a majority at Brussels. (Yes=23; No=1; Abstentions=2)
Postal votes: 20 votes (out of 39 WG members) were returned (Yes= 17; No= 1; Abstentions=2).
Secondary marker: Sigalia carpatica. This planktonic foraminiferan is widespread in the Mediterranean Region of the Tethys. It is associated with Inoceramus siccensis and Texanites in Tunisia. In north Spain (Navarra) it is very close to the lowest occurrence of Cladoceramus undulatoplicatus.
This secondary marker has been supported by a postal ballot (Yes=10; No=3; Abstentions=7).
Some voting members have emphasized usefulness of other planktonic foraminiferan (D. Herm) and the convenience of benthic form as secondary markers in Boreal Realm (L. Kopaevich).
Texanites (Texanites). Its first occurrence is below the lowest Cladoceramus undulatoplicatus. It has been cited in assemblage with inoceramids normaly regarded as Coniacian .
Dicarinella asymetrica. Its first appearance is above the lowest Sigalia carpatica, and therefore is not suitable to characterize the Coniacian-Santonian boundary. It is restricted to basinal facies.
Platyceramus siccensis. Only known from N. Africa, although it may be correlated with other areas through the lowest S. carpatica.
Sphenoceramus pachti. It is a boreal species rare at middle latitudes and unknown in palaeotropics (Tethys).![]()
As yet, we cannot make a formal proposal, because we need to know and integrate the biostratigraphy better.
We have selected three candidates:
1) Olazagutía Quarry (Navarra, Spain). Prof. Lamolda would collate data and report to the Chairman.
2) Seaford Head (Sussex, England). Dr. Mortimore and Mr. Wood would collate data and report to the Chairman.
3) Ten Mile Creek (Dallas, Texas). Prof. Kauffman and Dr. Gale would collate data and report to the Chairman.
This proposal was approved UNANIMOUSLY. (Yes=34; No=0; Abstentions=0)
The postal ballot also supported this porposal (Yes=18, No=1, Abstentions=1).
Dr. J. Salaj have sent a new proposal for the C/S boundary stratotype in the El Kef Area, in Tunisia, whose fossil content is cited above elsewhere. Ch. Colom has sent a proposal for this boundary in the Boreal Province; several sections are located at the Smoky River, Bad Heart River and Tipper's Coulee, all of them in west-central Alberta (Canada), having a very rich fossil content. Also, Dr. L. Kopaevich has proposed a section at Emdy-Kurgan, Eastern Boreal Europe, which is being studied by Naidin, Kopaevich and Gale.
Criteria for the subdivision of the Santonian Stage have been changeable since De Groussouvre (1901) proposed two zones in the Corbières, a lower Santonian Mortoniceras texanum Zone and an upper Santonian Placentyceras syrtale Zone. Both indices are American species and unknown in France (Hancock, 1991). In Europe, both Texanites gallicus and Texanites quinquenodosus are common in the lower Santonian, but they are not recorded in the Corbières (Kennedy and Wright, 1983; see above Ammonite chapter). For the upper Santonian several indices have been used, thus Placenticeras paraplanum Wiedmann seems to characterize top Santonian (Hancock, 1991; Kennedy, 1995), whereas in N. Spain the index could be Eupachydiscus isculensis (Redtenbacher) (Martínez et al., 1996).
Results outside Europe are also confusing and show endemic zonations, whose correlations up till now are not suitable. It is possible to distinguish two to five subdivisions, according to the literature.
Inoceramids show a similar picture. A detailed zonation inthe N. German-Polish Basin (Tröger, 1989) is not recognized everywhere (see Inoceramid chapter for lowermost Santonian Tröger's Zone 25). Thus, in N. Spain (Martínez et al., 1996) 4 or 5 subdivisions are recognisable, but most of them are quite different from Tröger's zones, because boreal taxa - mainly the genus Sphenoceramus - have not been found in Spain; except for the lower Santonian Cladoceramus undulatoplicatus Zone, which is very common worldwide. Kennedy (1995) suggested the first occurrence of the cosmopolitan species Cordiceramus cordiformis (Sowerby) as the base of the Middle Santonian.
Kennedy (1995) favoured the first appearance of the cosmopolitan Uintacrinus socialis Grinnell as the base of the Upper Santonian (see also Gale et al., 1995). Nevertheless Leahy and Lerbekmo (1995) have shown that the Western Interior appearance of crinoid Uintacrinus socialis (and Marsupites) should be time transgresive relative to its appearance in Europe, according with its correlation to the base of Chron 33r.
The Santonian of N.W. Europe is subdivided into five Gonioteuthis zones, whereas the Santonian of the Russian Platform is subidivided into only two zones (Christensen, 1990). Nevertheless, the base of the Santonian cannot be defined on the basis of belemnites, because Gonioteuthis is extremely rare in the lower Lower Santonian. The L.O. of Cladoceramus undulatoplicatus may be identical or very close to the base of the coranguinum/wesfalica Zone, and Gonioteuthis granulata (Blainville) appears slightly later than Uintacrinus socialis (Christensen, written comm.). Russian workers favoured a two fold division of the Santonian, in boreal regions (Kopaevich, Yazykova, written comm.).
Recent research on planktonic foraminifera has changed known occurrences (Robaszynski et al., 1984) for both Globotruncanita elevata (Brotzen) and Globotruncanita stuartiformis (Dalbiez). The first occurrence of G. elevata is younger (see Premoli-Silva & Sliter, 1994, fig. 2) and probably suitable to correlate the basal 'upper' Santonian. Both Dr. Salaj and Dr. Kopaevich disagreed with Santonian occurrences of Globotruncana arca (Cushman), because those younger specimens actually belong to ancestral species in the phylogeny of G. arca. Lamolda supports that opinion: he used to find (unpublished data) Globotruncana convexa Sandidge in north Spain 'upper' Santonian rocks, but not G. arca s.s. First occurrences of G. convexa and Globotruncana manaurensis Gandolfi had been cited by Sigal (1977) as upper Santonian markers in the Mediterranean Region (also, see Salaj, 1980). Status and ranges of above mentionned species should be cleared but they seem interesting for Santonian subdivision.
Nannofossils present several problems related to preservation of the genera Calculites and Lucianorhabdus. Nevertheles, Calculites obscurus (Deflandre) Prins & Sissingh used to be a marker of 'upper' Santonian and Lucianorhabdus cayeuxii Deflandre for 'middle' Santonian (Wagreich, 1992). The first occurrence of Amphizygus minimus Bukry was reported by Burnett (1996) close to the first occurrence of Uintacrinus socialis Grinnell. In the Bottaccione Section, Gardin et al. (submitted) have found a similar sequence of nannofossil events - successive occurrences of L. cayeuxii followed by A. minimus - to those in England (Gardin, written comm.). Therefore, it will be necessary to check other sections to contrast correlations of these indices, especially the relative horizons of L. cayeuxii, A. minimus, U. socialis, and C. obscurus.
After discussion this motion was put:
1) A 3 fold division of the Santonian;
2) Formal proposals are premature at present;
3) A possible datum for the base of the Middle Santonian is the extinction point of Cladoceramus undulatoplicatus.
4) A possible datum for the base of the Upper Santonian is the first occurrence of Uintacrinus socialis.
The WG voted, at Brussels: Yes=15; No=0; Abstentions=6.
The postal ballot supported the motion by a mayority: Yes=12, No=5, Abstentions=3)
At present, the WG has no proposals for the substage boundary stratopypes. Possible boundary sections in Texas, along the Channel Coast, in Germany and Mangyshlak were discussed. Integration of macro-, micro-, nannofossil datum-levels, stable isotope-ratios and palaeomagnetism, is necessary before further progress is possible.
This report is the result of the collaboration of many members of the Santonian Working Group. The collaboration of J. M. Hancock in the final production of this report has been especially useful.![]()
Bailey, H.W., Gale, A.S., Mortimore, R.N., Swiecicki, A. & Wood, Ch.J., 1983. The Coniacian-Maastrichtian Stages of the United Kingdom, with particular reference to southern England. Newsletters on Stratigraphy, 12: 29-42.
Bailey, H.W., Gale, A.S., Mortimore, R.N., Swiecicki, A. & Wood, Ch.J., 1984. Biostratigraphical criteria for the recognition of the Coniacian to Maastrichtian stage boundaries in the Chalk of north-west Europe, with particular reference to southern England. Bulletin geological Society of Denmark, 33: 31-39.
Birkelund, T., Hancock, J.M., Hart, M.B., Rowson, P.F., Remane, J., Robaszynski, F., Schmid, F. & Surlyk, F., 1984. Cretaceous stage boundaries-Proposals. Bulletin geological Society of Denmark, 33: 3-20.
Burnett, J., 1996. Nannofossils and Upper Cretaceous (sub-)stage boundaries - state of the art. Journal of Nannoplankton research, 18: (in press)
Christensen, W.K. 1990. Upper Cretaceous belemnite stratigraphy of Europe. Cretaceous Research, 11: 371-386.
Coquand, H., 1857. Position des Ostrea columba et biauriculata dans le groupe de la craie inférieure. Bulletin Societé géologique de France, 2(4): 745-766.
Dhondt, A.V., 1992. Cretaceous inoceramid biogeography: a review. Palaeogeography, Palaeoclimatology, Palaeoecology, 92: 217-232.
Dobrovs, S.A. & Pavlova, M.M., 1959. Inoceramus. In: Moskvin, M.M. (Editor), Atlas verkhnemelovoi fauny severnogo Kavkaza i Kryma. Gosudartsvennoe Nauchnotekhnicheskoe Izdatel'stvo Neftyanoj i Gornotoplinnoj literarury. Moskva, pp.130-163.
Flores, J.A., Gorostidi, A. & Lamolda, M.A., 1987. Nanoflora y bioestratigrafía del paso Coniaciense-Santoniense en Alava noroccidental. Paleontologia i Evolució, 20(1986): 151-163.
Gale, A.S., Montgomery, P., Kennedy, W.J., Hancock, J.M., Burnett, J.A. & McArthur, J.M., 1995. Definition and global correlation of the Santonian-Campanian boundary. Terra Nova, 7: 611-622.
Gardin, S., Del Panta, F., Monechi, S. & Pozzi, M., Toward a better definition of the Santonian/Campanian and Campanian/Maastrichtian boundaries. New calcareous nannofossil results from the Bottaccione section (Gubbio, Italy). (submitted)
Grossouvre, A. de, 1901. Recherches sur la craie supérieure. I. Statigraphie générale. Mémoires Societé Géologique de France, pp. 1-1013.
Hancock, J.M., 1991. Ammonite scales for the Cretaceous System. Cretaceous Research, 12: 259-291.
Herm, D., Kauffman, E.G. & Wiedmann, J., 1979. The age and depositonal environment of the 'Gosau' Group (Coniacian-Santonian), Brandenberg/Tirol, Austria. Mitteilungen der Bayerischen Staatssamlung für Paläontologie und historiche Geologie, 19: 27-92.
Jarvis, I. & Gale, A.S., 1984. The Late Cretaceous transgression in the S.W. Anglo-Paris Basin: stratigraphy of the Craie de Villedieu Formation. Cretaceous Research, 5: 195-224.
Kauffman, E. G., Sageman, B. B., Kirkland, J. I., Elder, W. P., Harries, P. J., & Villamil, T., 1993. Molluscan biostratigraphy of the Cretaceous Western Interior Basin, north America. In: Caldwell, W.G.E. & Kauffman, E.G. (Editors), Evolution of the Western Interior Foreland Basin. Geological Association of Canada, Special Publication, 39: 397-434
Kennedy, W.J., 1995. Defining the base of the Santonian and its substages using macrofossils. Report to the Santonian Working Group.
Kennedy, W.J., Bilotte, M. & Melchior, P., in press. Ammonite faunas, biostratigraphy and sequence stratigraphy of the Coniacian-Santonian of the Corbières (N.E. Pyrenees). Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine.
Kennedy, W.J. & Wright, C.W., 1983. Ammonites polyopsis Dujardin, 1837 and the Cretaceous ammonite family Placentyceratidae Hyatt, 1900. Palaeontology, 26: 855-873.
Larson, P.A., Morin,R.W., Kauffman, E.G. & Larson, A., 1991. Sequence stratigraphy and cyclicity of Lower Austin/Upper Eagle Ford outcrops (Turonian-Coniacian), Dallas County, Texas. Guidebook, DGS Field Trip 9. Dallas Geological Society, Austin, 61 pp.
Leahy, G.D. & Lerbekmo, J.F., 1995. Macrofossil magnetobio- stratigraphy for the upper Santonian-lower Campanian interval in the Western Interior of North America: comparisons with European stage boundaries and planktonic foraminiferal zonal boundaries. Canadian Journal of Earth Sciences, 32: 247-260.
López, G., Martínez, R. & Lamolda, M.A. 1992., Biogeographic relationships of the Coniacian and Santonian inoceramid bivalves of northern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology, 92: 249-261.
Martinez,R., Lamolda, M.A., Gorostidi, A., López, G. & Santamaría, R., 1996. Bioestratigrafía integrada del Cretácico Superior de la Región Vascocantábrica. Revista española de Paleontología, nº extraordinario: 157-168.
Master, B.A., 1977. Mesozoic planktonic foraminifera, a world-wide review and analysis. In: Ramsay, A.T.S. (Editor), Oceanic Micropaleontology,1. London. Academic Press, pp. 301-731.
Moskvin, M.M., (Editor), 1986. Stratigraphiya SSSR. Melovaya sistema Akademia Nauk SSSR, 1, 338 pp.
Premoli-Silva, I. & Sliter,W.V., 1994. Cretaceous planktonic foraminiferal biostratigraphy and evolutionary trends from the Bottaccione section, Gubbio, Italy. Palaeontographica Italica, 82: 1-89.
Robaszynski, F., Caron, M., González Donoso, J.M. & Wonders, A.H. (eds.) & European Working Group on Planktonic Foraminifera, 1984. Atlas of Late Cretaceous Globotruncanids. Revue de Micropaléontologie, 26: 145-305.
Salaj, J., 1975. Contribution à la microbiostratigraphie du Mésozoïque et du Tertiaire de la Tunisie septentrionale. Ve Colloque Afriquain Micropaléontologie, Addis Ababba. Revista española de Micropaleontología. Monografías: 703-783.
Salaj, J., 1980. Microbiostratigraphie du Crétacé et du Paléogène de la Tunisie septentrionale et orientale (Hypostratotypes tunisiens). Geologicky Ústav Dionyza Stúra, Bratislava, 238 pp.
Seitz, O., 1961. Die Inoceramen des Santon von Nordwestdeutschland. I. Teil (Die Untergattungen Platyceramus, Cladoceramus und Cordiceramus). Beihefte zum Geologischen Jahrbuch, 46: 1-186.
Seitz, O., 1965. Die Inoceramen des Santon und Unter-Campan von Nordwestdeutschland. II Teil (Biometrie, Dimorphismus und Stratigraphie der Untergattung Sphenoceramus J. Böhm). Beihefte zum Geologischen Jahrbuch, 69: 1-194.
Seitz, O., 1970. Über einige Inoceramen aus der Oberen Kreide. 1. Die Gruppe des Inoceramus subquadratus Schlüter. 2. Die Muntigler Inoceramenfauna und ihre Verbreitung im Ober-Campan und Maastricht. Beihefte zum Geologischen Jahrbuch, 86: 1-171.
Sigal, J., 1952. Aperçu stratigraphique sur la micropaléontologie du Crétacé. XXe Congress Géologique International, Alger. Monographies régionales (1), 26, 45 pp.
Sigal, J., 1977. Essai de zonation du Crétacé méditerranéen à l'aide des foraminifères planctoniques. Géologie Méditerranéenne, 4: 99-108.
Sissingh, W., 1977. Biostratigraphy of Cretaceous calcareous nannoplankton. Geologie en Mijnbouw, 56: 37-65.
Toshimitsu,S., Matsumoto, T., Noda, M., Nishida, T. & Maiya, S., 1995. Towards an integrated mega-, micro- and magneto-stratigraphy of the Upper Cretaceous in Japan. Journal Geological Society of Japan, 101:19-29 (in japanese)
Tröger, K.-A., 1989. Problems of Upper Cretaceous Inoceramid Biostratigraphy and Palaeobiogeography in Europe and Western Asia. In: WIEDMANN, J. (Editor), Cretaceous of the Western Tethys. Schweizerbart'sche Verlagsbuchhandlung, Stuttgart, pp. 911-930.
Tröger, K.-A. & Summerberger, H., 1994. Coniacian and Santonian inoceramid bivalves froom the Gosau-Group (Cretaceous, Austria) and their biostratigraphic and palaeobiogeographic significance. Annual Naturhistorische Museum Wien, 69A: 161-197.
Ueda, Y., 1962. The Type Himenoura Group, with Paleontological Notes by T. Matsumoto & Y. Ueda. Memoirs Faculty of Science, Kyushu University, ser. D., 12(2): 129-178.
Vasilenko, V.P., 1961. Upper Cretaceous Foraminifera from the Mangyshlak peninsula. Trudy VNIGRI 171.
Wagreich, M., 1992. Correlation of Late Cretaceous calcareous nannofossil zones with ammonite zones and planktonic Foraminirera: the Austrian Gosau sections. Cretaceous Research, 13: 505-516.
Yazykova, E.A., 1996. Post-crisis recovery of Campanian desmocertacean ammonites from Sakhalin, Far East Russia. In: Hart, M.B.(Editor), Biotic Recovery from Mass Extinction Events. Geological Society Special Publication, 102: 299-307.
Young, K.,1963. Upper Cretaceous ammonites from the Gulf Coast of the United States. University of Texas Publication, 6304: 1-373.
![]()
![]()
{kind=link}